| トップ | 教室紹介 | 研究概要 | メンバー | 研究発表 | 募集 |
教授 佐藤 元彦 (Motohiko SATO, MD, PhD)
略歴
| 1987年 | 旭川医科大学医学部医学科卒 |
| 1987年-1993年 | 旭川医科大学第一内科 |
| 1994年-1996年 | 米国サウスカロライナ州立医科大学 Postdoctoral Fellow |
| 1997年-2001年 | 旭川医科大学医学部 助手 |
| 2001年-2006年 | 米国ルイジアナ州立大学医学部 Assistant Professor |
| 2006年-2012年 | 横浜市立大学大学院医学研究科循環制御医学 准教授 |
| 2012年3月-現在 | 愛知医科大学医学部生理学講座 教授 |
学位
1993年 医学博士 旭川医科大学
研究内容
- 疾病に関連する新規G蛋白活性制御因子の同定
-
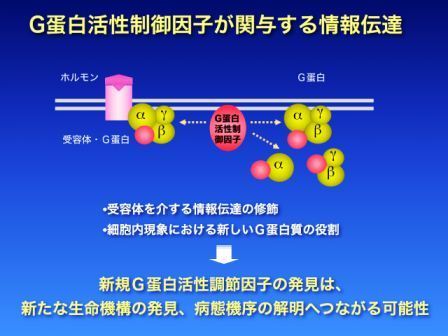
ヒト疾病の発症・進展に三量体G蛋白は重要な役割を果たします。細胞膜のG蛋白共役受容体はホルモンにより刺激を受けると三量体G蛋白質を活性化し、活性化されたG蛋白質が下流分子に情報を伝えます。つまり、三量体G蛋白質は細胞外のホルモン刺激を細胞内へ伝える重要な分子スイッチとして働きます。
しかし近年、三量体G蛋白は受容体以外の蛋白(G蛋白活性調節因子)によって活性化されることが解ってきました。G蛋白活性調節因子は三量体G蛋白をコントロールし、多彩な細胞現象に関与することが明らかになってきています。例えば、G蛋白の一種であるGαiサブユニットを制御する蛋白は細胞内輸送、オートファジー、また神経の軸索伸展に関与することが報告されています。G蛋白活性調節因子は、それぞれ独自のシグナル経路を形成して機能調節を行うと考えられています。このような調節機序はG蛋白活性調節因子が研究されるまで認識されていなかった機構です。
私たちは酵母を用いた機能的スクリーンシステムを用いて、これまで10種類のG蛋白活性調節因子を同定してきました。この方法を応用することで、病気で発現するG蛋白活性調節因子、すなわち病態調節を行うG蛋白活性調節因子を同定することが可能です。実際、私たちは虚血心モデルと心肥大モデルより、それぞれの病態に関与する新規G蛋白活性調節因子(AGS8、およびAGS11-13)を同定し、その病態への関与を報告しました。現在も病態、生命現象に関与する新たなG蛋白活性調節因子の同定が進行中です。
-
- シグナル調節因子を標的とした新しい心筋保護法の開発
-
狭心症、心筋梗塞をはじめとする虚血性心疾患の治療は循環器疾患の中でも非常に重要な分野です。心筋虚血の発生には交感神経をはじめとするG蛋白シグナルも重要な役割を果たします。
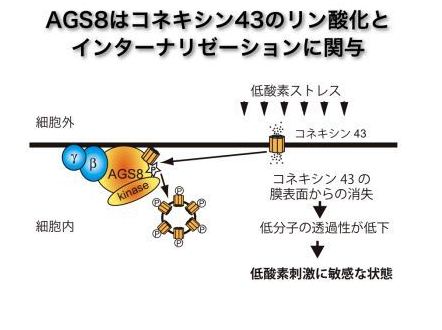
我々はラット狭心症モデルより、虚血心筋でG蛋白シグナルを制御する蛋白(Activator of G-protein signaling 8、AGS8)を同定しました。AGS8は低酸素により発現が上昇し、虚血心筋でG蛋白質(Gβγ)シグナルを制御していました。AGS8をノックダウンしたところ心筋細胞の低酸素誘導アポトーシスは著明に抑制され、AGS8-Gβγ経路が虚血心筋傷害に重要であることがわかりました。その後の検討から、AGS8-G蛋白複合体はチャネル蛋白(コネキシン43)の透過性を調節し細胞死に関与していると考えられました。
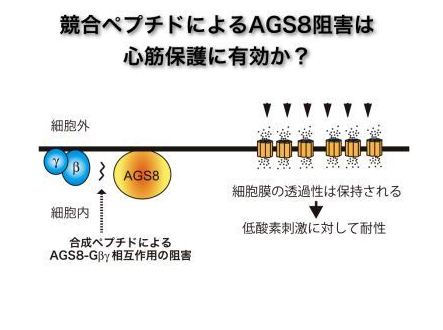
そこで我々は、AGS8によって活性化されるGβγシグナルを阻害することにより、心筋細胞を低酸素誘導細胞死から保護することを試みています。すなわち、AGS8のG蛋白が結合する部分を解析し、そのアミノ酸配列より結合を阻害するペプチドを合成しました。現在、このペプチドにより低酸素での細胞死を予防できるか、新しい心筋保護法になるか検討しているところです。
-
- 核内G蛋白が関与する転写調節機構の解明
-
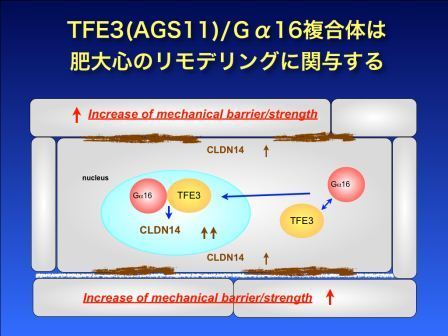
心臓の異常により心臓の筋肉が厚くなってくること(心肥大)があります。心肥大は心臓の機能異常の始まりとして重要な病態です。 私たちは、心肥大に関与するG蛋白活性調節因子を探索し、最近、マウス心肥大モデルより3種類の新規G蛋白活性制御因子(Activator of G protein Signaling 11-13, AGS11-13)を同定しました。これら3つの蛋白は同一ファミリーに属する転写因子であり、そのうち1つは遺伝子改変マウスの解析から心肥大に関与する可能性が報告されていたものです。
私たちの検討結果は、G蛋白活性制御因子でもある転写因子がG蛋白の活性と局在に影響を与えることを示していました。G蛋白活性制御因子(AGS11)はG蛋白を細胞核内に移行させ特定遺伝子の転写を著しく亢進させました。これは、肥大心のリモデルリングの新たな転写調節機構の存在を示唆していました。これまでこれら蛋白とG蛋白の相互作用の報告はなく、新規性に富む興味深い知見を示せたと考えています。現在、その詳細な機序を検討しています。
-
担当科目
生理学、生理学実習、胸部ユニット(循環器学、呼吸器学、アレルギー学)、総合医学
代表論文
- Activator of G-protein signaling 8 is involved in VEGF-induced choroidal neovascularization.
Hayashi H, Mamun AA, Takeyama M, Yamamura A, Zako M, Yagasaki R, Nakahara T, Kamei M, Sato M.
Scientific Reports 9(1): 1560, 2019. -
Activator of G-protein signaling 8 is involved in VEGF-mediated signal processing in angiogenesis.
Hayashi H, Mamun AA, Sakima M, Sato M.
Journal of Cell Science 129(6): 1210-1222, 2016. -
Protection of cardiomyocytes from the hypoxia-mediated injury by a peptide targeting the activator of G-protein signaling 8.
Sato M, Hiraoka M, Suzuki H, Sakima M, Mamun AA, Yamane Y, Fujita T, Yokoyama U, Okumura S, Ishikawa Y.
PLoS One 9(3):e91980, 2014. -
Roles of accessory proteins for heterotrimeric G-protein in the development of cardiovascular diseases.
Sato M.
Circulation Journal 77(10): 2455-2461, 2013. -
Identification of transcription factor E3 (TFE3) as a receptor-independent activator of Gα16: gene regulation by nuclear Gα subunit and its activator.
Sato M, Hiraoka M, Suzuki H, Bai Y, Kurotani R, Yokoyama U, Okumura S, Cismowski MJ, Lanier SM, Ishikawa Y.
Journal of Biological Chemistry 286(20): 17766-17776, 2011. -
Activator of G protein signaling 8 (AGS8) is required for hypoxia-induced apoptosis of cardiomyocytes: role of G betagamma and connexin 43 (CX43).
Sato M, Jiao Q, Honda T, Kurotani R, Toyota E, Okumura S, Takeya T, Minamisawa S, Lanier SM, Ishikawa Y.
Journal of Biological Chemistry 284(45): 31431-40, 2009. -
Identification of a receptor-independent activator of G protein signaling (AGS8) in ischemic heart and its interaction with Gbetagamma.
Sato M, Cismowski MJ, Toyota E, Smrcka AV, Lucchesi PA, Chilian WM, Lanier SM.
Proc Natl Acad Sci USA 103(3): 797-802, 2006. -
Accessory proteins for G proteins: partners in signaling.
Sato M, Blumer JB, Simon V, Lanier SM.
Annual Review of Pharmacology and Toxicology 46: 151-187, 2006.